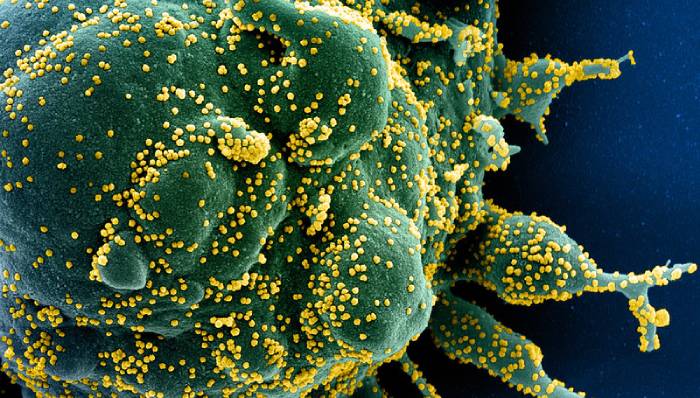
Imagine cu celulă infectată cu noul coronavirus (virusul, în galben, văzut la microscop electronic)
Cel mai frecvent, infecţia omului cu SARS-CoV-2 (coronavirus de tip 2 producător al sindromului acut respirator sever; agentul infecțios al pandemiei care a început în decembrie 2019 în provincia Wuhan din China) se produce prin inhalare pe cale respiratorie sau prin contaminare (prin „mâini murdare” pe cale fecal-orală) la nivelul mucoasei bucale. Virusul se mai poate transmite prin salivă, lichide biologice sau prin contactul rănilor deschise tegumentare cu suprafeţe contaminate.
Pe calea cea mai frecventă, la nivelul mucoasei nazale sau bucale, virusul se cuplează cu receptorii ACE2 (ACE2 = enzima de conversie a angiotensinei II) localizaţi pe suprafaţa externă a membranelor celulelor epiteliale de la acest nivel.
Receptorii ACE2 sunt distribuiți în numeroase țesuturi și organe: mucoasa nazală, mucoasa bucală, nazofaringe, plămâni, rinichi, inimă, endoteliul și pereții vaselor de sânge din stomac, intestin subțire, colon și creier, în mucoasa duodenului, jejunului, ileonului și în pereții musculari ai acestora, în piele și în glandele anexe ale acesteia (1). Tot în mucoasa bucală și nazală sunt prezente în număr redus și ușor variabil interindividual, specific fiecărui individ uman, celule ale sistemului imunitar de tipul macrofagelor, celulelor dendritice, limfocitelor și celulelor natural killer, care supraveghează aceste teritorii de potențiale agresiuni infecțioase.
Odată intrat în celula epitelială, virusul SARS-CoV-2 eliberează ARN-ul viral în citoplasma acesteia, unde se folosește de ribozomii celulei pentru a traduce mai întâi parțial informația genomului viral în proteine virale cu rol enzimatic (replicaze și proteaze) de care virusul se va folosi pentru a multiplica genomul viral și învelișurile proteice virale, respectiv nucleocapsida și anvelopa.
Virusul induce un efect patogen direct asupra celulei infectate prin parazitarea compartimentului intracelular de sinteza proteică, astfel că resursele celulare energetice (ATP-ul) și materia primă – aminoacizi și glucide - sunt deturnate către producția de virus.
Noile particule virale se asamblează intracelular, apoi prin exocitoză ies, posibil cu alterarea integrității celulei-gazdă, invadând alte celule care exprimă receptorul ACE2. În acest timp celulele infectate reacționează prezentând pe suprafața externă a membranei celulare proteine virale (cu rol de antigen) cuplate cu MHC-I, (MHC I = complexul major de histocompatibilitate I). MHC I e prezent pe toate celulele nucleate din organism și este particular pentru fiecare individ, funcționând ca un cod molecular de identificarea a celulelor ca fiind proprii/ale organismului și nu ale altui organism.
Celulele rezidente local ale sistemului imunitar (rezidente pentru a le diferenția de cele care circulă prin vasele de sânge) scanează suprafața membranară externă a celulelor epiteliale în căutarea de antigene. Într-un organism competent imunitar, prezența antigenului non-self în combinație cu MHC I pe suprafața celulei reprezintă un semnal ca respectiva celulă să fie atacată de limfocitele T citotoxice, membrana ei perforată de către enzimele eliberate de acestea sau de către sistemul complement și toate componentele celulei infectate să fie degradate enzimatic de către macrofage.
Când este însă un agent infecțios nou, cu care organismul nu s-a mai întâlnit, lucrurile nu mai sunt la fel de simple, ci este necesară captarea particulelor virale libere de către niște celule specializate anume pe captarea, prelucrarea și prezentarea antigenelor. Aceste celule specializate se numesc celule prezentatoare de antigen (CPA) și sunt prezente în cantitate variabilă în majoritatea țesuturilor, mai ales în cele care reprezintă o barieră fizică față de mediul extern. Reprezentanții principali ai CPA sunt macrofagele și celulele dendritice. Ele înglobează particula virală într-o veziculă pe care o endocitează, o digeră parțial prin proteoliză într-un proteasom, eliberează în spațiul extracelular niște citochine pentru a chema întăriri (citochinele pot fi metaforic asemănate cu niște rachete de semnalizare intercelulară, chemoatractanți pentru alte celule imunitare), apoi migrează în ganglionii limfatici regionali unde prezintă fragmentele proteice rezultante mici (de până la 10 aminoacizi) din particula endocitată, în combinație cu molecule ale CMH II (CMH II = complexul major de histocompatibilitate II), pe suprafața externă a membranei celulare către limfocitele T helper și către limfocitele B în regiuni ganglionare dedicate acestei interacțiuni. Moleculele de CHM II au și ele o structură caracteristică fiecărui individ, însă sunt mai multe forme de CMH II la același individ, fiecare cu afinitate ușor diferită de legare pentru diverse antigene, ceea ce duce la o variabilitate interindividuală a promptitudinii și competenței răspunsului imunitar la stimulii antigenici. Moleculele de CMH II diferă atât prin afinitatea de legare a antigenului, cât și ca densitate pe suprafața celulei, în dinamică ducând și la o variabilitate temporală intraindividuală a competenței imunitare.
Limfocitele B au atașate pe suprafața externă a membranei niște imunoglobuline cu structura de anticorpi. Atunci când la întâlnirea cu CPA (celula prezentatoare de antigen) vine un limfocit B care are pe membrană imunoglobuline (anticorpi) complementare spațial și chimic cu antigenul prezentat, complementaritate pe sistemul „cheie-în-broască”, limfocitul se activează, secretă o multitudine de citochine pentru a recruta ajutoare (alte limfociteT și B), se multiplică (expansiune clonală autolimitată) și apoi începe să sintetizeze imunoglobuline (anticorpi) similare cu cele atașate membranei celulare, pe cei nou-sintetizați îi va elibera însă în circulație în concentrație din ce în ce mai mare. Astfel vor apărea în sânge anticorpi liberi să detecteze atât virusurile libere, cât și pe cele expuse de către celulele infectate. Anticorpii au forma literei Y, cu extremitățile brațelor recunosc antigenul, cu zona de ramificație activând complementul, iar cu coada se pot lega de macrofage și de celulele natural killer. Această capacitate a anticorpului de activa complementul și de a se lega cu coada Y-ului de macrofage sau celulele natural killer se manifestă numai după ce anticorpul a găsit antigenul și s-au cuplat. Astfel că numai după legarea antigenului conformația anticorpului se schimbă discret, exteriorizându-se niște situsuri active care vor declanșa pasul următor al apărării imunitare, adică fagocitarea și eliminarea complexelor anticorp-antigen sau eliminarea celulelor infectate.
Complexul antigen-anticorp este fagocitat, lizat și eliminat în principal de către macrofage. Complementul e un complex proteic care în mod normal e dezasamblat, iar componentele lui circulă inactive și dizolvate în plasmă. În inflamație aceste componente se activează enzimatic succesiv una pe cealaltă, se numește activare în cascadă, și se combină până când se întregește complexul numit sistem complement, care arată ca un sistem de tuburi ce perforează membrana celulei-țintă, căreia îi crește permeabilitatea, iar celula-țintă se va dezintegra din cauza pierderii controlului schimbului de electroliți și al presiunii hidrostatice crescute în urma pătrunderii lichidului extracelular în interiorul celulei. Sistemul complement reprezintă un instrument de apărare al imunității nespecifice, numită și înnăscută în timp ce limfocitele T si B sunt reprezentanții imunității dobândite numită și adaptativă. Deoarece anticorpii au proprietatea de a activa complementul, separarea dintre imunitatea adaptativă și înnăscută este doar teoretică, ele completându-se reciproc pentru îndeplinirea obiectivului de eliminare a agentului infecțios.
Limfocitele T helper preiau antigenul de la CPA și îl prezintă la rândul lor către limfocitele T citotoxice până când antigenul va fi recunoscut de către receptorul de suprafață TCR (TCR = T cell receptor) al unui limfocit T citotoxic (se numește citotoxic deoarece poate neutraliza celulele infectate). Limfocitul T citotoxic, care este complementar spațial și chimic antigenului prezentat, se va lega de acesta prin TCR, apoi limfocitul T se activează, secretă citochine și se multiplică (expansiune clonală autolimitată) urmând a circula pentru recunoașterea și eliminarea celulelor-țintă. După seria de activare și multiplicare, limfocitele T devin sensibile la apoptoză.
Apoptoza este o moarte celulară programată care poate fi declanșată din interiorul celulei însăși sub presiunea a diverși factori cum sunt stresul oxidativ prelungit, acumularea de produși toxici, denaturarea proteinelor ori mutații genetice. Organismul oprește multiplicarea limfocitelor citotoxice specifice unui anumit antigen atunci când diminuează cantitatea de citochine, semn indirect că agentul prim declanșator ai inflamației, antigenul, a început sa scadă.
Un aspect necunoscut sau puțin investigat până în momentul de față este numărul de multiplicări permise limfocitului T citotoxic. O ipoteză neverificată ar fi aceea că ar putea exista un nivel de epuizare a potențialului mitogen a acestora odată cu înaintarea în vârstă, știind că există mecanisme genetice și epigenetice care limitează numărul de multiplicări ale tuturor celulelor din organism, așa cum este scurtarea telomerelor după fiecare diviziune celulară mitotică.
Din momentul pătrunderii virusului până la eliminarea lui din organism, multipli factori celulari și plasmatici se coordonează reciproc pentru îndeplinirea obiectivului. În literatura de specialitate se întâlnește adeseori termenul de orchestrare, fiind necesară menținerea proporțiilor și integritatea fiecărui element participant pentru a evita complicațiile sau eșecul răspunsului imunitar.
Consecutiv infecției virale cu SARS-CoV-2, numeroase citochine sunt eliberate în circulație de către celulele imunitare implicate (macrofage, celule dendritice, celule natural killer, limfocitele T helper și citotoxice si limfocitele B), unele articole pe acest subiect menționează: interleukina 1 (IL 1), IL 2, IL 6, IL8, interferon 1, interferon 2, IL 10, IP 10 (proteina 10 indusa de interferon), factorul alfa de necroză tumorală, chemokine CCL2, CCL3, CCL5, CCL10 (numele de CCL vine de la structura moleculara ce conține punți de legătură cisteină-cisteină) (2,3). Mare parte dintre acestea sunt implicate predominant în faza de la infectare până la prezentarea antigenelor de către CPA către limfocite și numai dacă ceva nu merge bine în orchestrarea răspunsului imunitar nivelurile crescute de citochine persistă, dând o stare de hiper-inflamație sau furtună citochinică „cytokine storm” observată adeseori în formele fatale de boală.
Sunt descrise mai multe forme de interferon 1 (aproximativ 14), deoarece se cunosc 13 variante de gene care codifică interferonul alfa, care este principalul reprezentant al interferonului de tip 1 (IFN 1) și interferonul beta. IFN 1 mai cuprinde interferonul epsilon, omega, etc (probabil ca se vor mai descoperi și altele care apar numai în anumite condiții). Sunt grupate în clasa de IFN 1, deoarece se leagă la aceiași receptori. Multiplele forme de IFN 1 au efecte biologice parțial diferite ca urmare atât a afinității variabile de legare, cât și cascadei de evenimente intracelulare consecutive internalizării receptorului cuplat cu IFN 1 și procesării lui intracelulare (4). Se crede că nu toate efectele legării IFN1 sunt cunoscute. De-a lungul timpului s-a observat ca întrucât organismele infectate viral dezvoltă un răspuns inflamator în care INF 1 are rol central, virusurile au acumulat gene ce codifică proteine care inhibă sinteza de interferon -1 în organismele gazdă fie la nivel genetic prin blocarea promoterului genei care codifică IFN 1 (promoterul e o secventa de ADN situată la începutul genei, esențială pentru declanșarea transcrierii genei din ADN in ARN mesager și ulterior în proteină), fie prin blocarea directă a activității IFN 1, acest mecanism fiind mai greu de depistat și mai greu de intervenit pe el (5).
Pentru SARS-CoV și MERS-CoV s-a demonstrat scăderea efectelor IFN-1 epigenetic prin modificarea histonelor (pentru MERS-CoV) prin inhibarea semnalizării intracelulare consecutive legării IFN 1. La studierea genomului SARS-CoV-2 la 103 pacienți infectați s-a observat că zona de gene cu cea mai mare rata a mutațiilor este în regiunea ORF8 (open reading frame 8, adică a 8-a secvență de genom viral cuprinsă între un codon start și unul stop, din care rezultă o proteină mare care ulterior este fragmentată de către proteinaza virală în diverse proteine mai mici; 8 pentru ca este după al 8-lea codon start, urmând convenția de citire de la capătul 5’ al catenei ARN) (5). Se poate presupune că virusul are nevoie numai de întârzierea eliberării de IFN-1, până când încărcătura virală a consolidat teritoriul cucerit, fiind tot mai greu de eliminat știind că organismul trebuie sa elimine propriile celule infectate și că eliminarea virusului ar echivala cu o victorie a la Pirus.
Alt studiu (6) a incubat într-o linie celulară mai întâi unele gene ale virusului SARS-CoV-2: ORF6, ORF8 și codificatoarea proteinei N (a nucleocapsidei), apoi genele pentru interferon 1 uman și un control negativ, adica doar cu genele IFN 1. În linia celulară care a primit și genele virale, sinteza de IFN 1 a fost scăzută. De aici s-a tras concluzia ca proteinele virale codificate de ORF 8, ORF 6 și N inhibă sinteza de IFN-1. Mai mult decât atât, inhibă și transcrierea genelor care ar fi în mod normal activate de către IFN-1 (6).
Punând cap la cap studiile 5 și 6 se observă că cea mai susceptibilă zonă din genomul viral pentru mutații, ORF 8, are rol în inhibarea secreției de IFN 1, rezultând o variabilitate a răspunsului imunitar al gazdei în funcție de gradul de inhibare a IFN-1 de către virus, timpul de reacție al organismului (deoarece organismul uman își modifică strategia de apărare imunitară odată cu înaintarea în vârstă), ceilalți efectori ai răspunsului imunitar și capacitatea fiziologică de adaptare a funcțiilor uzuale: circulație, respirație, contracție miocardică, filtrare glomerulară renală, sinteză hepatica de proteine, înnoire a celulelor imunitare, etc la stresul nou-apărut, atunci când resursele corpului trebuie judicios împărțite atât pentru eliminarea agentului infecțios cât și pentru menținerea funcțiilor vitale. Pentru un organ vital deja suferind, un stres nou și prelungit poate fi fatal.
Un lucru mai puțin înțeles este cum inhibă virusul celulele sistemului imunitar producătoare de IFN 1, odată ce el infectează numai celulele cu receptori ACE2 pe suprafață care sunt neimunitare. Din această cauză este necesară elucidarea traseului virusului SARS-CoV-2 după infectare și celulele care îl procesează, neputând exclude posibilitatea ca după internalizarea virusului, unele CPA-uri cu o capacitate limitată de izolare a virusului în proteasom și cu bagaj enzimatic de procesare redus să scape virusul de sub control și acesta să inhibe eliberarea de IFN-1 și să se multiplice nestingherit.
La persoanele care au avut forme ușoare de boală după infecția cu SARS-CoV-2, după vindecare s-a observat prezența de anticorpi antivirali din clasa IgG, neutralizanți față de proteina N (nucleocapsidă) și anticorpi specifici IgG pentru domeniul de legare al proteinei spike (RBD-S: receptor binding domain of S (spike) protein). De asemenea, persoanele testate au și limfocite T specifice pentru aceleași antigene, limfocite care au fost depistate prin capacitatea de eliberare a interferonului 2 (IFN2 = interferon gama) după ce au fost puse în contact cu respectivele antigene virale purificate 90% (7).
Un aspect relevant pentru rezistența la infecție sau vulnerabilitate este cantitate de limfocite circulante, urmărite în dinamică, după declanșarea răspunsului imun. La persoanele cu formă gravă de boală și mai ales la cele care au decedat, limfocitele au scăzut continuu (8, 9, 10 și 11). Sunt unele discrepanțe între studii privind severitatea bolii și limfopenia (limfopenie = număr scăzut de limfocite în sânge). De exemplu, autorii studiului 8 au găsit limfopenie la toate persoanele cu formă gravă de boală (69 de persoane), fără sa o detecteze la persoanele cu forme non-severe (135 de persoane) în timp ce autorii studiului 11 au găsit că limfocitele au scăzut la aproximativ 76% dintre persoanele infectate (29 din 39 de persoane incluse în studiu) deși numai 46% (18 din persoanele incluse în studiu) au avut formă severă de boală.
Deoarece sunt și rare persoane cu forme ușoare sau moderate de boală care se pot prezenta cu limfopenie, rezultă ca efectul limfopenic al infecției apare relativ precoce în evoluția bolii și deci precoce în organizarea răspunsului imunitar atunci când cooperarea intercelulară este maximă. Probabil că efectul limfopenic se corelează cu încărcătura virală, dar poate fi și o condiție preexistentă a imunităţii prin care limfocitele sunt mai susceptibile la apoptoză sub presiunea stresului inflamator. O întrebare neelucidată este dacă virusul induce prin efect direct moartea limfocitelor sau dacă este o buclă de acțiune cu mai mulți agenți moleculari mediatori ai inflamației prin care limfocitele T sunt eliminate. În mod normal, după orice infecție care solicită răspunsul limfocitelor T citotoxic, după eliminarea agentului infecțios, cea mai mare parte a limfocitelor T citotoxice specifice antigenului respectiv moare prin apoptoză și numai o mică parte persistă ca limfocite T de memorie (care au reacție rapidă de apărare în caz de reinfecție) .
Limfopenia persistentă consecutivă infecției virale, în formele grave de boală, pe lângă valoarea prognostică negativă sugerează abilitatea virusului de a interfera cu răspunsul imun și altfel decât strict antigenic. Știind însă că nu s-a demonstrat infectarea directă a limfocitelor de către virus, ipotezele care au fost emise pentru depleția limfocitară cuprind susceptibilitatea crescută la apoptoză a limfocitelor T într-un mediu celular cu concentrație crescută de citokine IL6 și TNF alfa (10) sau declanșarea apoptozei limfocitelor direct de către virus prin mecanisme încă neidentificate. Anticorpii au rolul de a lega și opsoniza particulele virale accesibile pe suprafața celulei sau în zona de tranzit intercelular a virusului de la o celulă la alta sau din circulație. Depozitul de virus intracelular însă, poate fi eliminat numai cu ajutorul limfocitelor T. Chiar dacă anticorpii pot mobiliza și sistemul complement care poate declanșa moartea celulelor opsonizate de anticorpi, dovezile de prezență constantă a limfocitelor T specifice de memorie post-infecție la majoritatea persoanelor care au depășit boala argumentează pentru rolul esențial al limfocitelor T pentru clearance-ul viral.
Un deziderat al cunoașterii patogeniei SARS-CoV-2 este cum anume este indusă limfopenia, deoarece prin combaterea acestui fenomen ar fi mai multe șanse pentru reechilibrarea răspunsului imunitar celular competent. Se presupune că moartea limfocitelor e produsă prin declanșarea apoptozei direct de către virus sau indirect prin dezechilibrul citochinic care ar sensibiliza limfocitele la apoptoza. Un aspect interesant este că spre deosebire de majoritatea celulelor din organism, limfocitele au o înclinație accentuată spre apoptoză, mai ales limfocitele implicate într-un răspuns imunitar acut și doar prezența unor citochine cu rol trofic inhibă calea mereu deschisă către apoptoză.
Organismele care nu pot elimina virusul, deși nu reușesc să monteze un răspuns imun consistent prin anticorpi și limfocite T citotoxice, hiperactivează partea de început a reacției imunitare, cu citochine. Citochinele în exces produc un status hiperinflamator și procoagulant în tot corpul și din aceste cauze pot apărea mici trombi în diferite organe, trombi care obstrucționează microcirculația. Organele cu microtrombi suferă de lipsa acută de oxigen și nutrienți, astfel că nu își mai pot îndeplini funcțiile și apar complicații amenințătoare de viață, în plămân – insuficiență respiratorie, în inimă – scad debitul cardiac și tensiunea arterială, risc de aritmii potențial fatale, în rinichi – se acumulează produși toxici în sânge, în ficat – se acumulează produși toxici, scade producția de glucoză, scade producția de proteine plasmatice care mențin compoziția și presiunea osmotica a sângelui normale, în intestin – crește permeabilitatea peretelui intestinal și risc de sepsis.
Odată cu înaintarea în vârstă se modifică strategia de apărare imunitară a organismului. De exemplu, celulele natural killer sunt mai multe, dar cantitatea de citochine și eficiența per celulă natural killer este diminuată, astfel creșterea cantitativă este o compensare a diminuării calitative. Consecutiv sunt necesare mai multe semnale intercelulare. Și cu macrofagele se întâmplă cam același lucru. Subtipurile de limfocite de asemenea își schimbă profilul cantitativ și calitativ. Se pare totuși că organismul uman deține informația pentru o îmbătrânire sănătoasă.
O posibilă corelație pentru răspunsul imunitar adeseori ineficient la organismele cu vârstă înaintată ar putea fi scăderea treptată a cooperării intercelulare. De exemplu, în recunoașterea antigenică a ARN-urilor virale sunt implicați receptori TLR 7 (12). Acești receptori odată activați, induc prompt eliberarea de interferon 1. Un studiu care a investigat densitatea receptorilor TLR pe celule dendritice plasmacitoide (aceste celule funcționează ca CPA care care preiau virusul din circulație și au cea mai potentă acțiune în eliberarea de interferon 1) la persoane cu vârstă medie 74 de ani vs. persoane cu vârstă medie 28 de ani a găsit ca prezența de TLR 7 și TLR 9 este mult scăzută pe celulele dendritice plasmacitoide de la seniori față de juniori (13). Același studiu susține o reducere de peste 60%, la seniori vs. juniori a proporției de celule dendritice plasmacitoide care pot elibera IFN 1 după stimularea cu virus gripal. Dealtfel, în populația americană >90% dintre pierderile anuale de vieți omenești din cauza infecției gripale apar la grupa de vârstă de peste 64 ani (14).
Ca o mențiune privind heterogenitatea tabloului clinic al bolii după infecția cu SARS-CoV-2, variabilitatea simptomelor e strâns legată de tropismul virusului, respectiv de celulele și organele pe care le poate infecta, de densitatea receptorilor ACE2 pe celulele respective, de imunitatea locală, cât și de cea a întregului organism. Deoarece tractul respirator, digestiv, rinichiul, inima, pielea, exprimă receptorul ACE2, simptomele de început ale infecției pot predomina într-unul dintre acestea, însă aspectul constant este ca în formele grave de boală, plămânii suferă cel mai mult.
Surse:
1. I. Hamming, et al. Tissue distribution of ace2 protein, the functional receptor for sars coronavirus. A first step in understanding sars pathogenesis J. Pathol.: A Journal of the Pathological Society of Great Britain and Ireland., 203 (2) (2004), p. 631e7 Google Scholar
2. Wagner Gouvea dos Santos, Natural history of COVID-19 and current knowledge on treatment therapeutic options, Biomedicine & Pharmacotherapy, Volume 129, 2020, 110493, ISSN 0753-3322, https://doi.org/10.1016/j.biopha.2020.110493.
3. Víctor J. Costela-Ruiz, Rebeca Illescas-Montes, Jose M. Puerta-Puerta, Concepción Ruiz, Lucia Melguizo-Rodríguez, SARS-CoV-2 infection: The role of cytokines in COVID-19 disease, Cytokine & Growth Factor Reviews, 2020, ISSN 1359-6101, https://doi.org/10.1016/j.cytogfr.2020.06.001.
4. Cherie T. Ng, Juan L. Mendoza, K. Christopher Garcia, Michael B.A. Oldstone, Alpha and Beta Type 1 Interferon Signaling: Passage for Diverse Biologic Outcomes, Cell, Volume 164, Issue 3, 2016, Pages 349-352, ISSN 0092-8674, https://doi.org/10.1016/j.cell.2015.12.027.
5. Xiaolu Tang, Changcheng Wu, Xiang Li, Yuhe Song, Xinmin Yao, Xinkai Wu, Yuange Duan, Hong Zhang, Yirong Wang, Zhaohui Qian, Jie Cui, Jian Lu, On the origin and continuing evolution of SARS-CoV-2, National Science Review, Volume 7, Issue 6, June 2020, Pages 1012–1023, https://doi.org/10.1093/nsr/nwaa036
6. Jin-Yan Li, Ce-Heng Liao, Qiong Wang, Yong-Jun Tan, Rui Luo, Ye Qiu, Xing-Yi Ge. The ORF6, ORF8 and nucleocapsid proteins of SARS-CoV-2 inhibit type I interferon signaling pathway,Virus Research,Volume 286,2020,198074,ISSN 0168-1702, https://doi.org/10.1016/j.virusres.2020.198074
7. Ling Ni, Fang Ye, Meng-Li Cheng, Yu Feng, Yong-Qiang Deng, Hui Zhao, Peng Wei, Jiwan Ge, Mengting Gou, Xiaoli Li, Lin Sun, Tianshu Cao, Pengzhi Wang, Chao Zhou, Rongrong Zhang, Peng Liang, Han Guo, Xinquan Wang, Cheng-Feng Qin, Fang Chen, Chen Dong, Detection of SARS-CoV-2-Specific Humoral and Cellular Immunity in COVID-19 Convalescent Individuals, Immunity, Volume 52, Issue 6, 2020, Pages 971-977.e3, ISSN 1074-7613, https://doi.org/10.1016/j.immuni.2020.04.023.
8. Ruyuan He, Zilong Lu, Lin Zhang, Tao Fan, Rui Xiong, Xiaokang Shen, Haojie Feng, Heng Meng, Weichen Lin, Wenyang Jiang, Qing Geng, The clinical course and its correlated immune status in COVID-19 pneumonia, Journal of Clinical Virology, Volume 127, 2020, 104361, ISSN 1386-6532, https://doi.org/10.1016/j.jcv.2020.104361.
9. Qianwen Zhao, Meng Meng, Rahul Kumar, Yinlian Wu, Jiaofeng Huang, Yunlei Deng, Zhiyuan Weng, Li Yang, Lymphopenia is associated with severe coronavirus disease 2019 (COVID-19) infections: A systemic review and meta-analysis, International Journal of Infectious Diseases, Volume 96, 2020, Pages 131-135, ISSN 1201-9712, https://doi.org/10.1016/j.ijid.2020.04.086.
10. Soheil Tavakolpour, Taha Rakhshandehroo, Erin X. Wei, Mohammad Rashidian, Lymphopenia during the COVID-19 infection: What it shows and what can be learned, Immunology Letters, Volume 225, 2020, Pages 31-32, ISSN 0165-2478, https://doi.org/10.1016/j.imlet.2020.06.013.
11. Z. Liu, W. Long, M. Tu, S. Chen, Y. Huang, S. Wang, W. Zhou, D. Chen, L. Zhou, M. Wang, M. Wu, Q. Huang, H. Xu, W. Zeng, L. GuoLymphocyte subset (CD4+, CD8+) counts reflect the severity of infection and predict the clinical outcomes in patients with COVID-19 J. Infect. (2020), Google Scholar
12. S.S. Diebold, T. Kaisho, H. Hemmi, S. Akira, C. Reis e Sousa Innate antiviral responses by means of TLR7-mediated recognition of single-stranded RNA Science, 303 (2004), pp. 1529-1531 Google Scholar
13. Jing Y, Shaheen E, Drake RR, Chen N, Gravenstein S, Deng Y. Aging is associated with a numerical and functional decline in plasmacytoid dendritic cells, whereas myeloid dendritic cells are relatively unaltered in human peripheral blood. Hum Immunol. 2009;70(10):777-784. doi:10.1016/j.humimm.2009.07.005 https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5718338/
14. Thompson WW, Shay DK, Weintraub E, et al. Mortality Associated With Influenza and Respiratory Syncytial Virus in the United States. JAMA. 2003;289(2):179–186. doi:10.1001/jama.289.2.179
15. Mihaescu Grigore. Imunologie si Imunochimie, Ed. Universitaţii din Bucureşti, 2001.
